|
Salt and
fresh waters are very different from each other on their optical properties. Optically
pure off-shore oceanic water absorbs shiefly red rays and transmits blue rays.
Therefore, oceans and open seas have typical blue color (similar to blue sky). Inland
seas are more turbid and have bluish colors with the transition to greenish ones.
Due to suspended particles fresh waters are much more turbid than in inland
seas. In general, turbid fresh water absorbs chiefly blue rays and transmits
red rays. Depending on the properties of suspended particles, fresh waters have
greenish, yellowish or even brownish (in peat bogs) colors. Only optically pure
fresh water in mountain lakes and drawn quarries is blue. 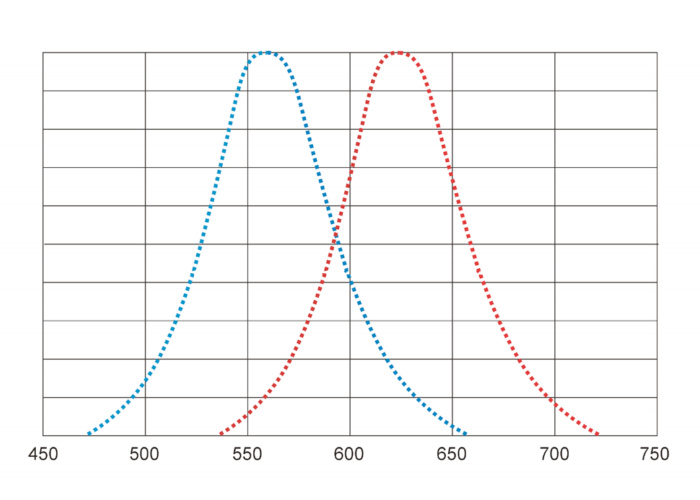
These differences in the optical mediums are reflected in
the spectral sensitivity of eyes in saltwater and freshwater fish. Spectral
sensitivity is an ability of the eye to perceive monochromatic light of equal
power with the different wavelengths. Eyes of saltwater fish, and human, are most sensitive to light with the
wavelengths of 550-560 nm (the green-yellow part of the spectrum) (the left
curve in the Fig.1). Reflecting optical properties of the freshwater medium,
eyes of freshwater fish are more sensitive to light with the wavelengths of
600-680 nm (the red part of the spectrum) (the right curve in the Fig.1). Due
to bell like dependence of spectral sensitivity equipower monochromatic light
of different wavelengths are not equally bright to the eye. Green light is most
bright for human and saltwter fish, red light is most bright for freshwater
fish contrary to our perception.
For
example, the maximum of spectral sensitivity in freshwater adapted threespined
stickleback, Gasterosteus aculeatus,
is near 605 nm (e.g., Rowe et al., 2004). In common carp, Cyprinus carpio, goldfish, Carassius auratus, and bluegill, Lepomis macrochirus, these maximums lie
in the region of 612-615 nm (Cronly-Dillon & Muntz, 1965; Tamura & Niwa, 1967). Grundfest (1932) gives the same results for
other Lepomis. In L. macrochirus, this maximum can be shifted to 620-640 nm (Hawryshyn et al., 1988). The maximum of
spectral sensitivity in largemouth bass, Micropterus
salmoides, which are the natural predators for the foregoing Lepomis, is shifted even to 673 nm (Kawamura
& Kishimoto, 2002). According to Matsumoto & Kawamura (2005), eyes of
common carp and Nile tilapia, Oreochromis niloticus, are sensitive to
light in the near-infrared part of the spectrum.
In
anadromous Far-Eastern redfin, Tribolodon
hakonensis, the maximum of spectral sensitivity is shifted from 548 nm, in
lake, to 612 nm, in more turbid pond (Kawamura & Kishimoto, 2002).
These features of spectral sensitivity in fish must be taken
into consideration in naturalistic, scientific, applied and other research.
Color vision of human is closer to that in saltwater fish. On the other hand, color vision in freshwater
fish and other freshwater animals, like tadpoles, is far different than ours.
Basic References
Cronly-Dillon
J.R., Muntz W.R.A. 1965. The spectral sensitivity of the goldfish and the
clawed toad tadpole under photopic conditions. Journal of
Experimental Biollogy 42, 481-493
Grundfest
H. 1932. The sensibility of the sun-fish, Lepomtis, to monochromatic radiation
of low intensities. Journal of General Physiology 15, 307-328
Hawryshyn
C.W., Arnold M.G., McFarland W.N., Loew E.R. 1988. Aspects of color vision in
bluegill sunfish (Lepomis macrochirus):
ecological and evolutionary relevance. Journal
of Comparative Physiology A164, 107-116
Kawamura
G., Kishimoto T. 2002. Color vision, accomodation and visual acuity in the
largemouth bass. Fisheries Science 68, 1041-1046
Matsumoto
T., Kawamura G. 2005. The eyes of the common carp an Nile
tilapia are sensitive to near-infrared. Fisheries
Science 71, 350-355
Rowe M.P.,
Baube C.L., Loew E.R., Phillips J.B. 2004. Optimal mechanisms fo finding and
selecting mates: how threespine stickleback (Gaserosteus aculeatus) should incode male throat colors. Journal of Comparative Physiology A190,
241-256
Tamura T.,
Niwa H. 1967. Spectral sensitivity and color vision of fish as indicated by
S-potential. Comparative Biochemistry
& Physiology 22, 745-754
|







 SUBSCRIBE
SUBSCRIBE