|
General
At present,
the most manufactures of artificial fishing lures offer spinners, in which
rotating blades and sinkers made usually in the form of body or head, are
combined together. Such the leadheaded spinners as Mepps Lusox and Abu Garcia
Morrum are typical examples. In this post, we will show that combinations of
two lures, made by dividing the leadheaded lures into heads and light spinners,
are much more effective.
Fig. 1 illustrates
scematically an integrated leadheaded lure and combinations of its two parts
connected with the short, middle and long line or steel leaders (usually up to
30-60 cm length).
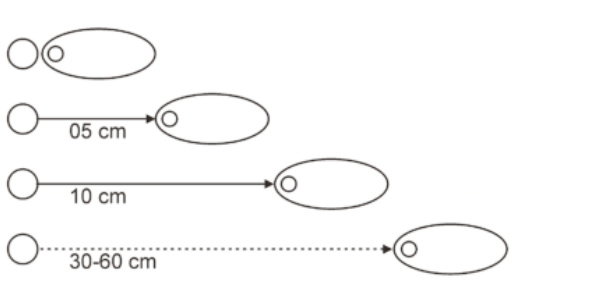
Figure 1. An integrated lure and tandems of its two parts
Field Experiments
We examined
the effectiveness of integrated lures (IL) in comparison with tandems of their two
parts, spherical heads and light spinners, connected with the 20 cm line leaders
(T). The high quality 5 gr stannic heads with the two lateral 2D or 3D eyes (5
mm, red, WTP Inc.), called goggle heads (gogglers), and Mepps Silver Aglia #2 were
used. Original Aglia spinners (see Mepps catalog) were disassembled and assembled
to replace the brass bodies of the hard plastic tubes (2.5 mm diameter, 20 mm
length, red).
In tests, goggle
heads were equipped with the small double hooks.
Using
common perch, Perca fluviatilis, as usable
model fish, lures of both types were presented alternately at the estimated
localities of these abundant fish. At each locality, 20 presentations (cast and
retrieving) of lures were made: 5 with IL, 5 with T, 5 with IL and 5 with T.
Then an experimentator moved to the other locality, where 20 presentations of
the compared lures were made in the reverse order. Within 1 day session, 39 perch were landed (and
released) in total, the numder of fish were distributed per earch 10 lure
presentations (for IL and T, correspondingly).
Student
t-test was used to estimate the difference between the mean values of fish
number for lures of both types, IL and T. Perch (71%) preferred tandems (p <
0,01). Among heads (about 2 cm length with hook) and spinners (4 cm), nine
perch of ten preferred to attack larger spinners.
Similar
results were obtained in the other 3 sessions with perch carried out in the
different fishing conditions. In one special test session (with the reassembled
Aglia Longue # 1), asp, Aspius aspius,
also authentically preferred spinners in tandems over single tinheaded lures.

Figure 2. Asp, Aspius aspius
It is necessary
to underline that strictly regulated test sessions have been conducted to
confirm the foregoing preferences at the statistical level. Meanwhile, the
members of our group use tandems of lures mentioned above and other (with
hookless leading lures, called usualy teasers)
during 50 years with the more or less constant seccess. The drawbacks of tandems
are the overlapping of leader for the main line, especially when lures are cast
upwind, and relatively short distance of casting.
Discussion
In an
experimental ethological literature, there is limited information about
responses of animals to two conjugately moving stimuli. Among two moving
feeding objects, edible frog, Rana
esculenta, for example, show preferences for near and larger objects (for
review, see Manteifel, 1977).
According
to tests and general observations in the nature, pursuits of horizontally
moving small objects, float up small objects and sinking small objects are
stereotypical motor acts in the feeding behaviour of fish (Protasov, 1968). These
motor acts as well as typical feeding poses play an important signaling role in
the intraspecific (like roach-roach) and interspecific (like perch-roach) relationships.
On the other hand, it is shown that pike, Esox
lucius, and other predatory fish prefer faster and discomposedly moving
feeding objects than benthivorous fish do (Meesters, 1940; Prorasov, 1968). In
our case, two lures allow to imitate all the foregoing patterns when an
evidently larger object pursues horizontally moving, lifting and sinking small
objects, using the well known lure slip, rise and fall retrieving technique.
Krause &
Godin (1996) exposed separately
nonforaging, horizontally foraging and nose-down foraging guppies, Poecilia reticulata, to an approaching cichlid fish predator model in aquaria.
Nonforaging guppies responded sooner to and initiated flight further away from
the approaching model than foraging fish
did collectively. At the same time, horizontally foraging individuals responded
sooner to the model than nose-down foraging ones. Comparing all test guppies,
nose-down foraging individuals were the most likely not to exhibit any response
to the predator model. Individual blue acara cichlid, Aequidens pulcher (6.5 cm), natural predators of guppies, preferred
to attack foraging prey over nonforaging ones and nose-down foraging prey over horizontally
foraging ones (2.0 cm all). An individual risk of predation for guppies
foraging nose-down was greater than for guppies foraging horizontally, and all
these fish were at greater risk than nonforaging guppies. This result was
consistent with the above differences in the guppy's responsiveness to an approaching
predation threat depending on their foraging behaviour. It is important in our
context that cichlid predators preferentially selected less wary and more vulnerable prey, that must be true for predators of
other species (references in Krause & Godin, 1996).
These data allow
to explain why predatory fish preferred in our experiments an overtake lure.
Basic References
Krause J. Godin
J-G.J. 1996. Influence
of prey foraging posture on flight behavior and predation risk: predators take advantage
of unwary prey. Behavioral Ecology 7, 264-271
Manteifel
Y.B. 1977. Visual system and behaviour in the anurans. Moscow, Science
Meesters A. 1940.
Ũber
die Organisation des Gesichtsfeldes der Fische. Zeitschrift für Tierpsychologie 4, 84-149
Protasov
V.R. 1968. Vision and near orientation in fishes. Moscow, Science
|







 SUBSCRIBE
SUBSCRIBE