|
General
Studying
behavioural responses of animals in experimental conditions, ethologists have
found relatively simple stimuli that could be more effective than the natural
objects. Single spots (called also eye-spots) and two horizontally arranged
spots, rectangular longitudinal stripes,
periodic gratings and other stimuli (Fig.1) belong to them. Because the foregoing
stimuli are not exact imitations of the natural objects, we will call them amimetic stimuli. In several articles,
we will group the main visual amimetic stimuli and describe how they are used
in the ethological research, whether they occur in the nature as well as their application
in the fishing lure industry.
In
the framework of applied ethology, we will address to the fishing lure
industry. It is that only sphere of the human activites, where artificial
stimuli and models of the various animals are used in the largest scale. 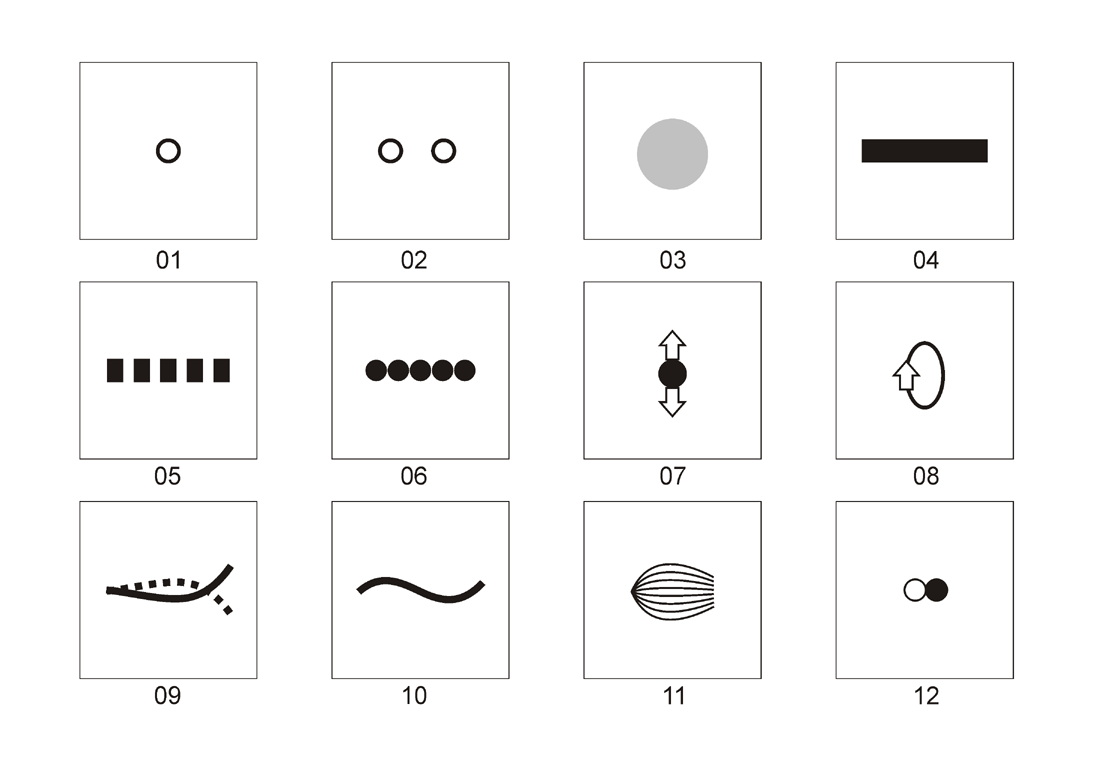
Figure
1. The basic visual amimetic stimuli
Names
of the basic visual amimetic stimuli used in this article:
01.
Concentric spots
02.
Two horizontal spots
03.
2D & 3D roundish stimuli
04.
Rectangular stripes
05.
Periodic gratings
06.
Chains
07.
Vibrators
08.
Spinners (rotating stimuli)
09.
Flutters
10.
Undulators
11.
Pulsators
12.
Mechanical & light flashers
Note,
stimuli 05 and 06 are periodic spatial, whereas stimuli 07, 08, 09, 10, 11, and
12 are periodic spatiotemporal
In
the terminology of early ethologists, some of the amimetic stimuli shown above called
the sign stimuli (e.g., Manning &
Dawkins, 1998). Generally, using simple stimuli and changing parts of complex
stimuli, scientists were able to find the so called supernormal stimuli that induced in animals the more strong
behavioural responses than the modelling natural objects.
For
example, the giant cane toads, Bufo
marinus, respond to the horizontally moving rectangular longitudinal stripes (20
mm long x 2.5 mm high) much more actively (on average of 10 times) than to live
crickets and insect plastic models (Robins & Rogers, 2004). Similarly,
reproductive males of the common toad, Bufo
bufo, prefer (in four cases against one) to form sexual pairs (Fig.2) with the
fixed blue discs (5 cm diameter) than with the live mobile females (Gnyubkin
& Kondrashev, 1978).

Figure 2. Reproductive males of toads prefer to congregate sexual pairs with blue discs than with live females
Manning
and Dawkins (1998) give many other examples of this kind.
Neuroethology
Visual
amimetic stimuli induce numerous behavioural responses in many animals and do
not imitate, as
mentioned above, the
concrete natural objects. The effectiveness of these stimuli is grounded on the common mechanisms of visual
perception, common for all visually guided animals. Among visual amimetic
stimuli, the nature of spots, stripes and gratings, both static and moving, as
well as rotating striped drums is most studied.
For example,
in fish and other
vertebrate animals, spots are detected at the level of ganglion cells of
retina, which have the more or less distinct concentric receptive fields with the
antagonistic center and periphery. According to Horn (1962; see Fig. 6.5 b,c), the boundaries of some spot are distinguished
depending on its size by one or several ganglion cells with the foregoing
receptive fields. Rectangular stripes and periodic gratings appeared in the
field of view are detected by the so called orientation selective ganglion
cells (e.g., Damjanović et. al., 2009). It is shown in numerous
electrophysiological tests that the cells of this type are represented by two
relatively distinct units called,
respectively, the detectors of horizontal lines and the detectors of vertical
lines.
In addition, on an example of developing larval
zebrafish, Brachydanio (Danio) rerio,
as an usable model object, neuromotor grounds of the behavioural responses to
artificial and natural visual stimuli are studied, in the larva development
(for review, see Portugues & Engert, 2009):
1) responses to large-filed moving
vertical gratings, called optomotor responses
2) responses to live parameciums and small
moving spots, called prey tracking
3) responses to large moving spots and
other large objects, called escape responses
Among other
mechanisms of adjusted effectiveness of amimetic visual stimuli matched with
the corresponding receptive fields, bilateral symmetry of spots and spatial
symmetry of gratings play an exceptionally inportant role (e.g., Kenward et
al., 2004). The effectiveness of pair stimuli is determined by the bilateral
symmetry of visual system and visual perception evolving during millions of
years in the field of the
Earth gravitation, but
the causes of the evolution of repetitive stilmuli and the corresponding
receptive fields are unclear. Kenward et al. (2004) consider about ten
factors that might lead to the evolution of repetitive visual stimuli and the
corresponding receptive fields, including the highest detectableness of
repetitive stimuli on the background of environmental optic noises.
In addition
to spatial amimetic visual stimuli, there are more complex spatiotemporal
amimetic visual stimuli (for review, see, e.g., Rothental, 2007). In
fish, responses to such spatiotemporal stimuli as rhythmic (temporal symmetric)
pulsations, vibrations and (worm-like) undulations are innate. In contrast to the spatial periodic gratings (Damjanović et al., 2009),
there are not distinct receptive units to detect these complex stimuli.
Basic References
Damjanović
I., Maximova E.M., Maximov V.V.
2009. On the organization of receptive fields of orientation-selective units
recorded in the fish tectum. Journal of Integrative Neuroscience 8, 323–344
Gnyubkin
V.F., Kondrashev C.L. 1978. Pair aggregation in the common toad, Bufo bufo L., in the reproductive
period. In: Mechanisms of animal vision. Moscow,
Science, 40-75
Horn
G. 1962. Some neural correlates of perception. Viewpoints in Biology.
Butterworth & Co. Publishers, London, p. 240-285
Kenward B.,
Wachtmeister C. A., Ghirlanda S., Enquist M. 2004. Spots and stripes: the
evolution of repetition in visual signal form. Journal of Theoretical Biology
230, 407-419
Manning
A., Dawkins M.S. 1998. An introduction to animal behaviour. 5th editon. UK, Camdridge
University Press
Portugues
R., Engert F. 2009. The neural basis of visual behaviors in the larval
zebrafish. Current Opinion in Neurobiology 19, 1–4
Robins A., Rogers L.J. 2004. Lateralized prey-catching responses
in the cane toad, Bufo marinus:
analysis of complex visual stimuli. Animal Behaviour 68, 567-575
Rothental G.G. 2007. Spatiotemporal dimensions of visual
signals in animal communication. Annual Review of Ecology, Evolution and
Systematics 38, 155–78
| 






 SUBSCRIBE
SUBSCRIBE