|
Kohda & Watanabe (1986, 1988) have
demonstrated that vertically striped fish, such as Coreoperca kawamebari and Macropodus
chinensis, prefer to rest at the vertically striped background. Of two horizontally
striped fish, Melanochromis auratus and Barbus titteya, M. auratus choose the horizontally striped
background, whereas B. titteya are
indifferent. At the same time, stripeless Acheilognathus limbata are slightly drawn towards the vertically
striped background, but stripeless Carassius
auratus show no preference (Kohda & Watanabe, 1988).
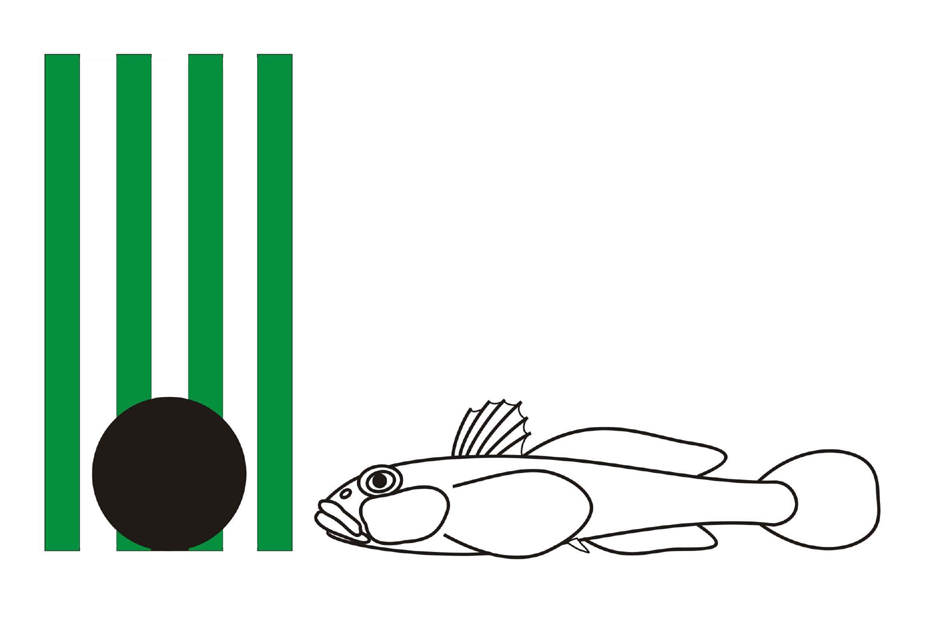
Fig.1 shows how stripeless round goby, Neogobius melanostomus, and other gobiids rest near the
vertically grating background with green stripes and an additional middle-size
black disc (20 mm diameter) attached at the foot of this background. If an
artificial background is sufficiently long (longer than 10-15 fish lengths),
gobies and other bottom fish, like common gudgeon, Gobius gobius, or common ruffe, Gymnocephalus cernuus, will rest usualy near the black disc (our
observations).
You can carry out own experiments attaching
the same or similar discs to the wall of your terrarium at the level of floor.
In these cases, small rodents will respond to these stimuli as an entrance to
the shelter. The same disc attached horizontally to the tip of vertical stick above
the water plays another function. Now all dragonflies and damselflies flying
beside will turn towards the disc to use it as an appropriate landing site.
Males of round and some other gobies
become black in the reproductive period (Meunier et al., 2009). In this period,
black discs or balls induce aggressive behaviuor in males and attract females
(our data). Reproductive red-belly males of three-spined stickleback, Gasterosteus aculeatus, respond to
various stimuli (Darkov, 1980). In particular, they display aggressive responses
to red geometric figures and avoid black ones.
Silvery lacustrine bleak, Leucaspius delineatus, aggregate with silvery
squares and silvery fish-like flat models (Darkov, 1980). They actively avoid black squares and other
black models, but slightly aggregate with live black molly, Poecilia sphenops.
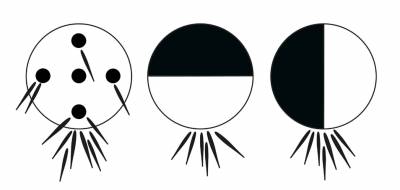
On the other hand, you can use black
discs or discs with the contrast edges and spots to bring together early fry of
mouthbreeding Mozambique
tilapia, Tilapia mossambica (Fig.2: Baerends,
1957).
When the
foregoing vertically striped background comes in motion, fish follow
more or less stable, depending on their ecology and locomotion, the moving background. In
general, this behaviour, named optomotor,
is innate, weakly dependent on the shape of visual stimuli (stripes or other
figures, see data by Shaw
& Sachs, 1967, for schooling fish, Menidia
menidia) and weakly
modified by the vital experience of animals. To study optomotor responses, fish are
usually placed into the circular transparent tanks that, in turn, are
surrounded with the rotating striped drums.
In fish, rotating striped drums are mainly used to study optomotor responses and large
field motion vision (e.g., Schaerer & Neumeyer, 1996).
Animals
demonstrate behavioural responses of other kinds in respect of small rotating
striped drums, small moving discs or other figures, small rotating sectorial or
dotted discs and flashing light spots.
In
particular, Magnus (1958) has showily used small rotating striped drums to
stimulte nuptial chases in males of butterfly silver-washed fritillary, Argynnis
paphia. Ishizawa (2005) has used striped cylinders and discs (6-18 cm in diameter) to study
hovering flight in golden-ringed dragonfly, Anotogaster
sieboldii.
In fish, small
moving and rotating stimuli are used to study small field
motion vision (Gehres & Neumeyer,
2007; Stojcev et al.,
2011). In conrast to responses to large field moving stimuli, responses to small field moving stimuli are dependent on the physiological state of animals and modified by their experience (Protasov, 1978). For example, fish learned to eat food of the certaine color will prefer small moving objects of the same or similar colors (due to the so called generalization).
These
examples together with other numerous findings, accumultated in experimental
ethology for decades, show that the same simple visual stimuli may induce
different behavioural responses in fish and other animals. It means that each
of these stimuli is not an imitation of some concrete object. To reflect this
irrefutable fact, stimuli of this kind are named non-simulative, or amimetic.
At this rate, the capability of amimetic stimuli to induce different
behavioural responses, depending on the context, must be grounded on
the common mechanisms of visual perception, common for all visually guided animals.
Basic References
Baerends
G.P. 1957. Behaviour: the ethological analysis of fish behaviour. In: The
physiology of fishes (ed. M.E.Brown). Vol.II, p.229-269, Academic Press Inc., New York Darkov A.A. 1980. Ecological features of visual signalization in fishes. Nauka, Moscow
Gehres M.M.,
Neumeyer C. 2007. Small
field motion detection in goldfish is red-green color blind and mediated by the
M-cone type. Visual Neuroscience 24,
399-407
Ishizawa N.
2005. The response to ratating objects by Anotogaster
sieboldii (Selys) males, Pt.2 (Anisoptera,
Cordulegastridae). Odonatologica
34, 211-218
Kohda Y.,
Watanabe M. 1986. Preference for vertical-striped backgrounds by the oyanirami Coreoperca kawamebari, a freshwater
serranid. Ethology 72, 185-190
Kohda Y., Watanabe M. 1988. Preference for
striped backgrounds by striped fishes. Zoological
Science 5, 501-503
Magnus D. 1958. Experimentelle
Untersuchungen zur Bionomie und Ethologie des Kaisermantels Argynnis paphia L.
(Lep. Nymph.): I. Über optische
Auslöser von Anfliegereaktionen und ihre Bedeutung für das Sichfinden der
Geschlechter. Zeitschrift für
Tierpsychologie 15,
397-426
Meunier B., Yavno S., Ahmed S. & Corkum L.D. 2009. First
documentation of spawning and nest guarding in the laboratory by the invasive fish,
the round goby (Noegobius melanostomus). Journal of Great. Lakes Research 35, 608-612 Protasov
V.R. 1978. Fish behaviour. The mechanisms of fish orientation and their use in
fishing. Food Industry,
Moscow
Schaerer S., Neumeyer C. 1996. Motion detection in goldfish investigated with the optomotor response is "color blind”. Vision Research 36, 4025–4034
Shaw E.,
Sachs B.D. 1967. Development of the optomotor response in the schooling fish, Menidia menidia. Journal of Comparative and Physiological Psychology 63, 385-388
Stojcev M.,
Radtke N., D'Amaro D., Dyer A.G., Neumeyer C. 2011. General principles in
motion vision: Color blindness of object motion
depends on pattern velocity in honeybee and goldfish. Visual Neuroscience 28, 361-370
| 






 SUBSCRIBE
SUBSCRIBE